
2.2.0
Ontogeny & Phylogeny
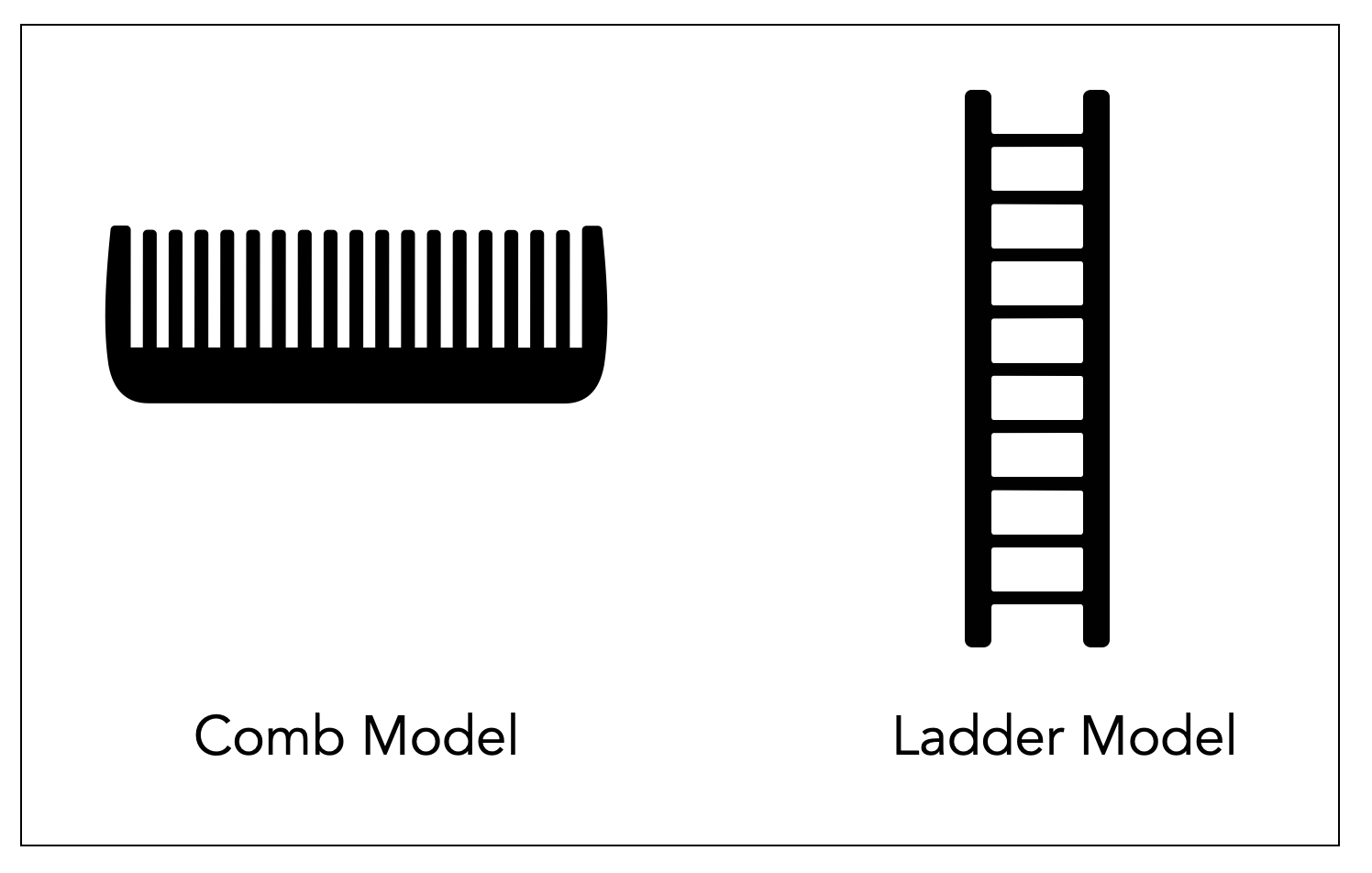
This chapter lays the groundwork for a later discussion of the PVT's hierarchical model. It defines terms such as ontogeny, phylogeny, and evolution and clarifies the debate on old beliefs. It also compares different views of evolution: staircase vs. cladistic (ladder vs. comb).
Ontogeny is you, from the moment of your conception to your birth — or later. It is the development of an individual human, animal, or plant. For humans and animals, it primarily refers to embryology.
Phylogeny studies a larger group, such as a phylum (we are chordates) or class (we are mammals). It relates to evolution.
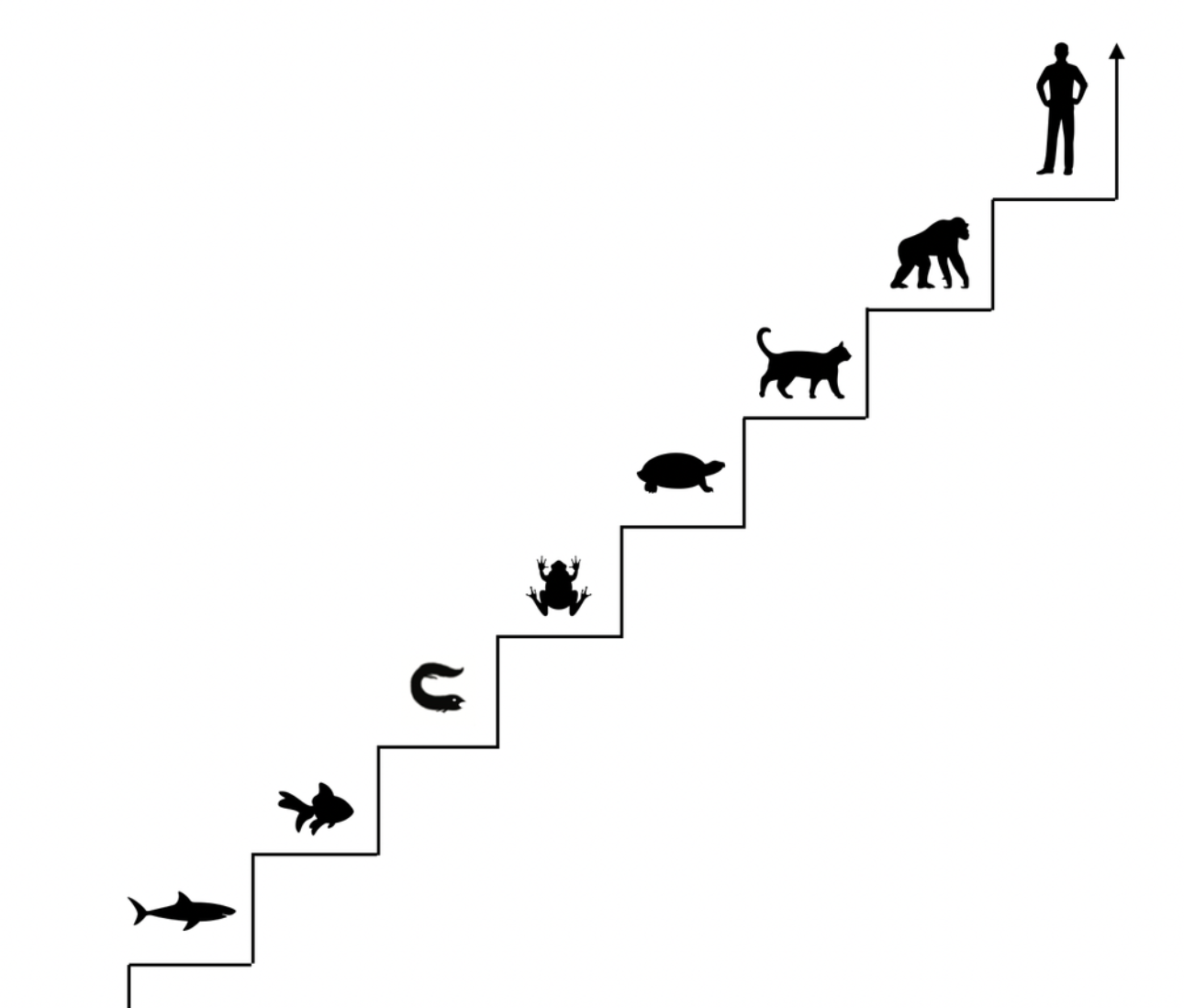
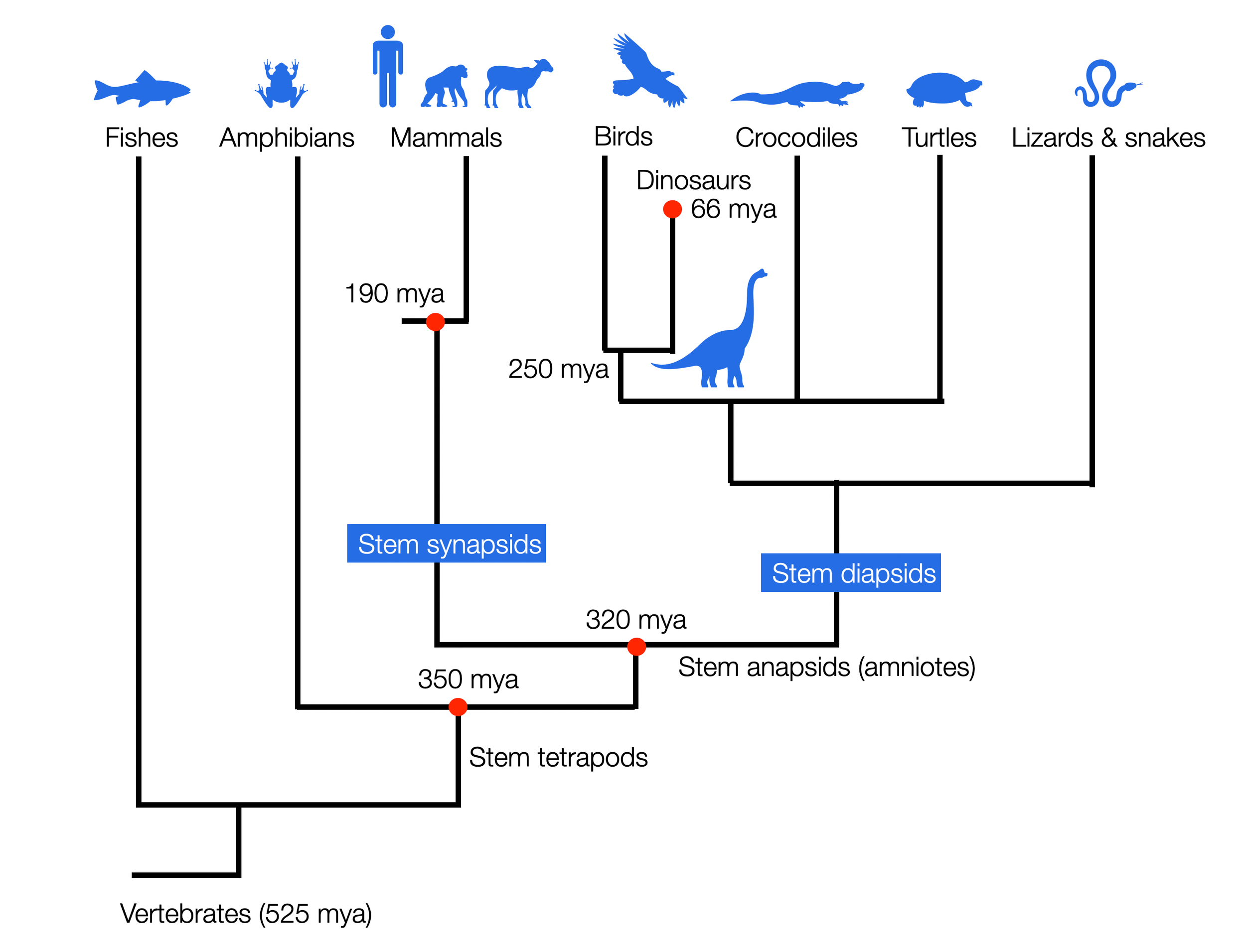
Comparison between the staircase perspective (Scala Naturae or The Ladder) and the cladistic perspective. Notice how , in the second graphic, humans stay on the same line as other species that survived. Graphic ©absb, 2024
Ontogeny vs. Phylogeny
Look at the diagram below! It strongly suggests that human development (ontogeny) and vertebrate development (phylogeny) follow the same direction — step by step — doesn't it? Becoming an adult is the logical end of human development. Any deviation from that path would have led you to the doctor. Is it the same with evolution? You can believe in evolutionary design or a polyvagal social agenda, but current science (especially biology) says otherwise.

Linking ontogeny and phylogeny is problematic because they radically differ in predictability. A strict genetic program at each stage guides the embryo's development into a fetus, baby, child, and adult. As a child, you could hardly stop yourself from growing taller and having a particular hair, eye, or skin color. “It's in my DNA,” as we sometimes say about our character or habits — or “it's in the family blood, you know!” If both parents are giants, their children will likely be the tallest in the class.
From a biological point of view, phylogeny is entirely different: it is random. Mutations don't occur according to a plan or design. New forms are selected if they have a better chance of survival. Sexual selection (hence the flowers' colors and the birds' parade) also plays a role.
The PVT doesn't see it that way. It implies that evolution has a social agenda so people can look at each other and feel calm and secure. A biologist would say this is as naive as believing that the banana is curved to fit our hand better or that the nose is designed to wear glasses. Well, biologists can be stubborn, can't they? That doesn't mean they're wrong.
Porges doesn't see the difference between psychology and biology or physiology (Berntson, 2007). He's obsessed with this mother-child visual exchange, which can be the sweetest thing in the world. The world doesn't stop there. Evolution cares little about feelings of security. It's all about survival. To do that, we need to maintain a healthy level of worry and fear. Is that not what makes mothers and fathers? Besides pride and joy, they are constantly worried about their children. They not only protect their outcomes, but try to optimize them, with healthy food, vitamins, or school support, breaking the limits of their genetic program.
Recapitulation
Recapitulation: Ontogeny Recapitulates Phylogeny
The "recapitulation" theory of evolution, also known as the "biogenetic law," proposes that the development of an organism from embryo to adult recapitulates or repeats the evolutionary history of its ancestors (e.g., fish, amphibians, reptiles).
The most famous proponent of recapitulation theory was the German biologist Ernst Haeckel in the late 19th century. He believed that the development of the embryo, from conception to birth (ontogeny), passes through stages that resemble or represent successive adult stages in the evolution of the animal's distant ancestors (phylogeny). It was first formulated in the 1820s by Étienne Serres, based on the work of Johann Friedrich Meckel.

Wikipedia Recapitulation. Haeckel’s drawings.
{kind=link}

This diagram illustrates the difference between Haeckel and von Baer. Haeckel proposed that embryos go through a developmental process that recapitulates adult stages, rather than transitional stages in which common patterns of organization are expressed (adapted from Wikipedia ©absb, 2024).
PVT and Recapitulation
What about the PVT? Although the PVT doesn't defend recapitulation per se, Porges, like Freud, postulates a link between the past and the present. Freud postulated that the father's murder happened in the distant past. Porges and Dana link our ups and downs to the steps of evolutionary phylogeny. Psychoanalysts think similarly when they discuss adult behavior as regressive-sometimes specifically to the oral, anal, or phallic stage. Drawing on Jacksonian dissolution theory, Porges provides a dramatic description of this regression (to use Freudian terms) from a civilized state of social engagement to immobility and collapse — through a state of struggle and flight. According to the PVT, responses to threats are phylogenetically inscribed in our nervous systems. In evolutionary retro-engineering, we go back down the evolutionary ladder every time we step outside the polyvagal framework of social engagement. In sum, recapitulation ascends the ladder, and dissolution (or regression for Freud) descends it.
Again, Porges doesn't advocate recapitulation — an outdated biological theory — but uses a similar concept: our personal history (ontogeny) and behavior (social or otherwise) reflect 500 million years of historical evolution. He sees a direct line from the first vertebrates to present-day humans, which contradicts the modern cladistic perspective.
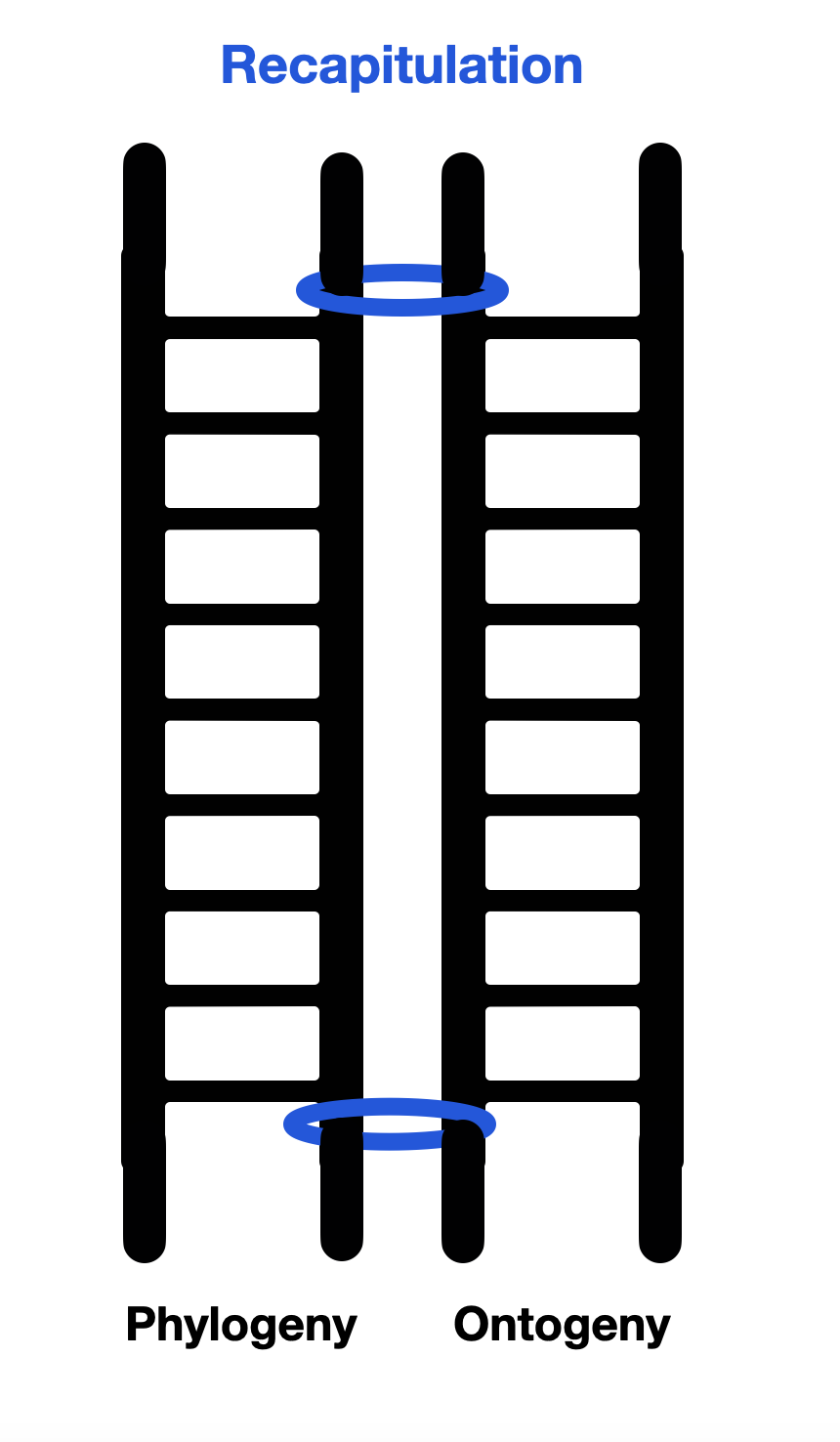
Diagram: Linking ontogeny and phylogeny, as the PVT does, is a slippery slope to recapitulation.
2.2.1
Old Biological Models
In part one (“Who?”), we saw how modern biology has discarded old models, such as the medieval ladder model. However, there are still popular models, such as MacLean's triune brain (which Peter Levine, Bessel van der Kolk, Pat Ogden, and Porges continue to promote and defend).
Recapitulation is another popular meme that has infected biology to this day. “Ontogeny recapitulates phylogeny,” also known as the biogenetic law or recapitulation theory, was proposed by German biologist Ernst Haeckel in the late 19th century. The theory suggests that the development of an individual organism (ontogeny) mirrors the evolutionary development of its species (phylogeny). As Stephen Jay Gould described in detail in Ontogeny and Phylogeny (1977), this theory is no longer valid in evolutionary science.
However, the PVT, and to a lesser extent, the Triune Brain Theory (MacLean), takes up the spirit of recapitulation by combining embryology and comparative anatomy. A good example is The Brain-Body Disconnect (Kearney, 2022), which attempts to pair the ontogeny and phylogeny of the vestibular system. In various articles, Porges presents his “1, 2, 3” hierarchy concept: one for the dorsal parasympathetic, two for the sympathetic, and three for the ventral parasympathetic. He argues that this order appears in embryonic development (ontogeny) and vertebrate evolution (phylogeny).

Building on Porges (2011), Dana (2018) proposes an evolutionary timeline that later serves to demonstrate the evolutionary nature of our responses to threat (e.g., immobilization or fight-flight). This is another form of recapitulation, as the PVT links phylogeny and physiology-two domains with very different timescales (millions of years vs. seconds or minutes). Using the Jacksonian term "dissolution" to describe our reactions to stress or threat is a form of reverse recapitulation. In this strange perspective, getting angry or paralyzed is an instant slide down the evolutionary ladder. The concept is exciting and easy to grasp. However, it's crucial to note that the current scientific literature doesn't support this whimsical concept. It's a reminder of the importance of aligning our theories with the latest research findings.
Before discussing more specific issues — such as the sympathetic or parasympathetic nervous system, which came first? — we need to understand some basic concepts.
2.2.2
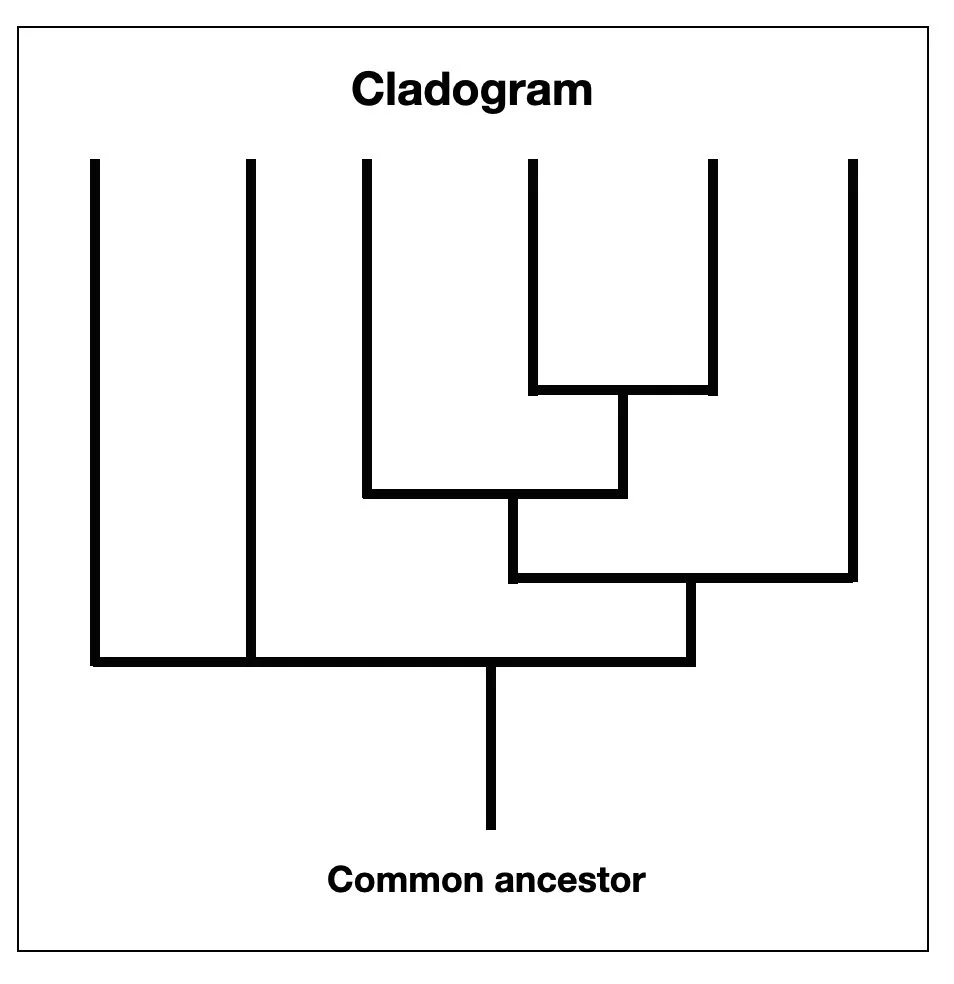
The cladistic revolution
The cladistic revolution began in the mid-20th century, mainly through the work of German entomologist Willi Hennig. In his seminal book, Phylogenetic Systematics (Hennig, 1966), he introduced the idea of grouping organisms based on:
common derived characters (e.g., four legs, lactation)
common ancestry.
His work marked a significant shift in how scientists approached the classification of species and the study of their evolutionary relationships. In the following years, cladistics became increasingly influential in systematics and taxonomy, significantly impacting our understanding of species relationships.
The cladistic revolution has profoundly impacted how we understand the relationships between species. Cladistics classifies species based on their common ancestry and shared evolutionary traits. This approach has shifted the focus from a linear hierarchical structure (a ladder), where species were viewed as more or less advanced, to a more nuanced understanding of the relationships between organisms. The image of a comb with equal-length indentations is more accurate. All living species, from ants to monkeys, are at the finish line of this million-year evolutionary race. It also replaces the traditional tree of life and places man at the pinnacle of creation.
In the context of the cladistic revolution, the terms “superior” and “inferior” become irrelevant because species are not ranked in a linear hierarchy. Instead, cladistics emphasizes that species are adapted to their specific ecological niches, and each organism is well suited to its particular environment. Similarly, “ancient” and “primitive” must be used cautiously.
Cladistics promotes a more holistic and egalitarian view of the natural world by focusing on a common evolutionary history and recognizing that all organisms are part of an intricate web of life. This approach challenges the anthropocentric perspective that places humans at the top of a hierarchy. It encourages a deeper understanding and appreciation of all life forms.

The cladistic revolution has replaced the medieval ladder, and Darwin's tree of life still contains an element of hierarchy that contradicts modern biology. In the three illustrations in this section, we use simple objects to symbolize the different perspectives (©absb, 2024).
It took us some time to understand where the paths diverge between the old models (Scala Naturae, Triune Brain, and PVT) and modern biology.
These three models are based on difference and exclusivity. They are elite models. Porges and MacLean are both crazy about reptiles that lack noble qualities. These inferior creatures have only one ear bone (instead of three) and no “smart vagus,” or they don't have a full mammalian brain.
Of course, we can't ignore the fact that Homo sapiens is now at the top of the food chain and even has the power to destroy the planet Earth. But before him, there was the era of bacteria, fish, dinosaurs, and stem mammals. We are only the next in line.
For us, MacLean and Porges don't fit the modern view of evolution, which is cladistic, not hierarchical. It's a comb model, not a ladder. In a comb, all dents are equally long-unless they are broken. As we will see in the various diagrams below, evolutionary paths are mostly parallel. Hagfish are not the ancestors of sharks, sharks are not the ancestors of fish, and reptiles are not the ancestors of us. We have common ancestors with them — with worms and flies. That's all. So, the polyvagal evolutionary timeline is primarily wrong.
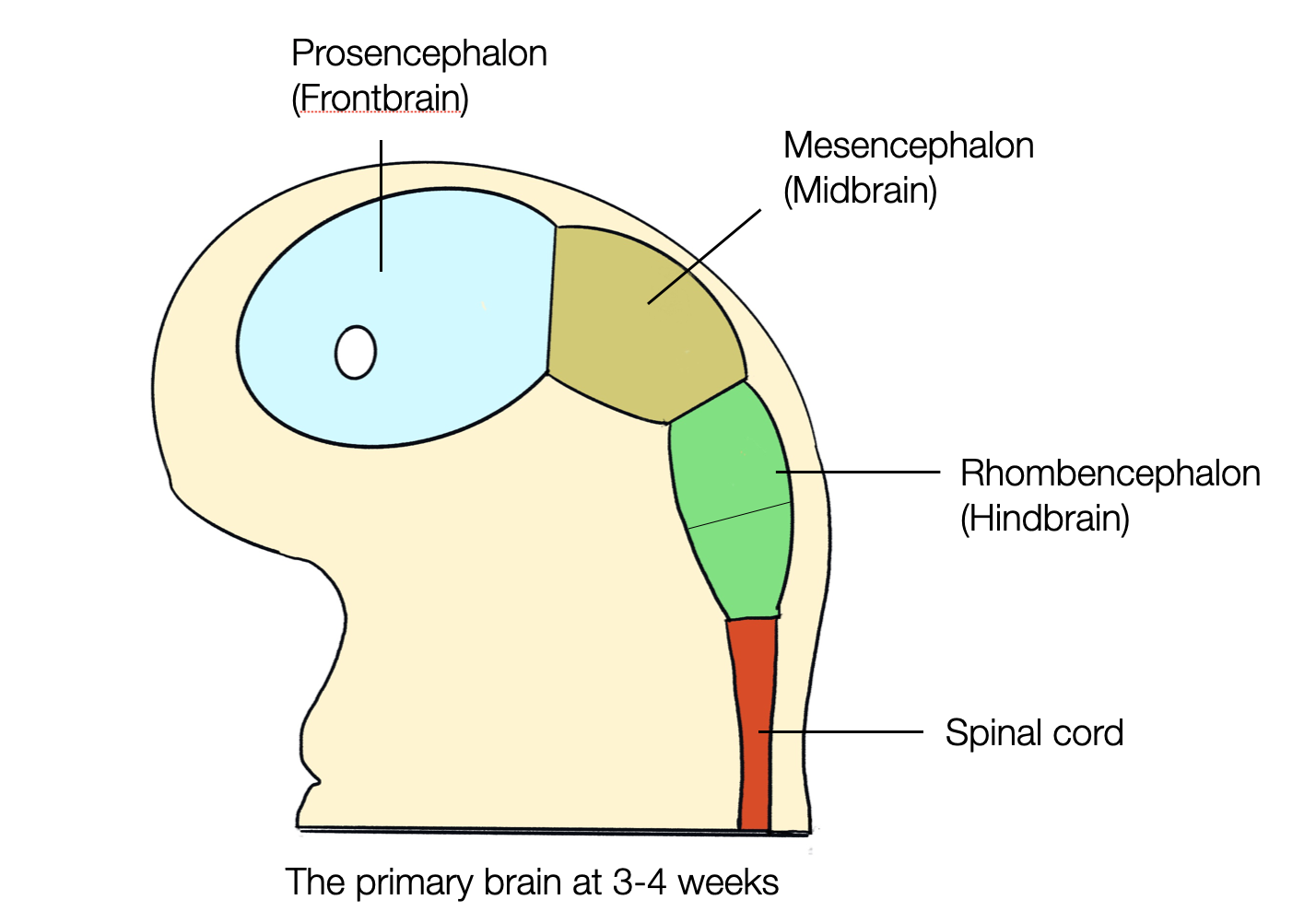
2.2.3
Since when has the brain three parts ?
The first bilaterians appeared 600 million years ago (mya). Although their brains were much smaller and simpler than ours, the basic tripartite organization (forebrain, midbrain, and hindbrain) was already present (see On the Origin and Evolution of the Tripartite Brain, Hirth, 2010). The regionalization of the head is based on a blueprint that has been conserved throughout evolution (see Towards a New Neuromorphology-Nieuwenhuys, Puelles, 2016). Therefore, the triune brain theory (MacLean) is invalid and outdated. (more about the blueprint in Hierarchy).
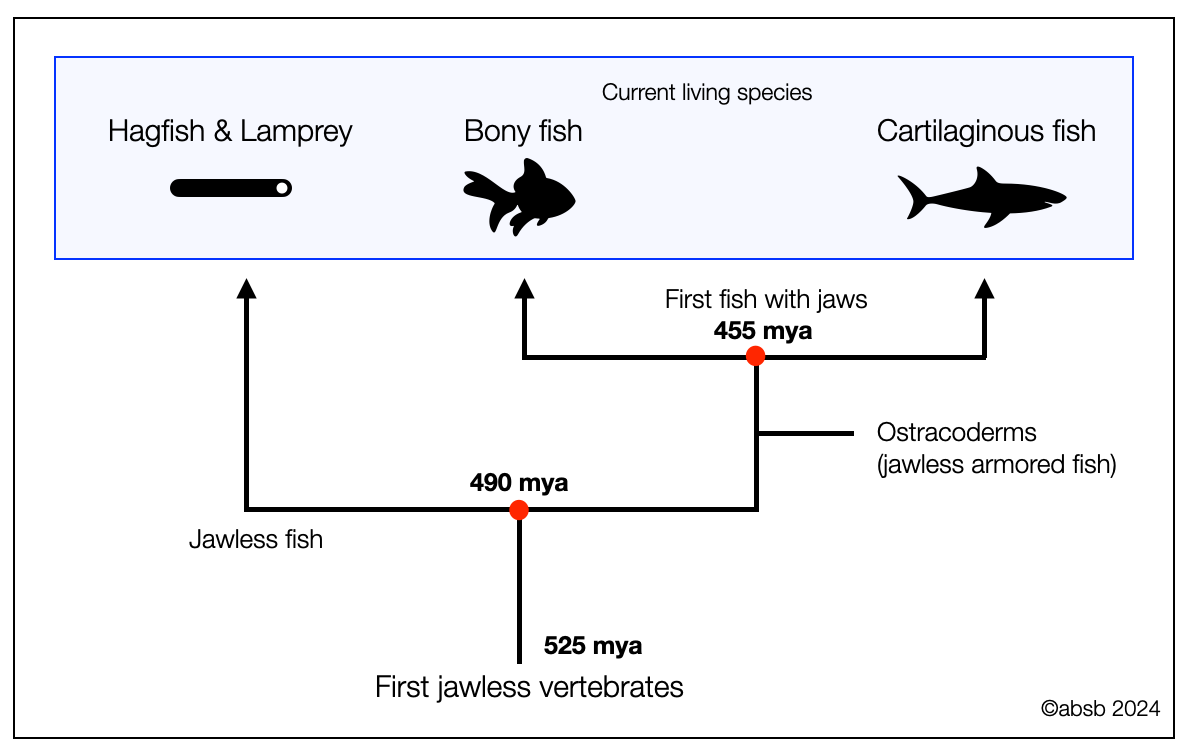
2.2.4
First Vertebrates
Vertebrates are animals (kingdom) belonging to the chordates (phylum), which have a notochord. The oldest vertebrates (about 500 mya) are called agnathans (Greek γναθοσ gnathos = jaw) because they don't have jaws. Because they disappeared long ago, we only know them from fossils, so we can't say much about their nervous systems.
However, we know more about modern jawless fishes that appeared later (200 mya), such as hagfishes and lampreys, which still exist today. They are called cyclostomes because of their round mouth (cyclostomes: κυκλοσ kuklos = circle and στομα stoma = mouth) bordered by dangerous teeth. Cyclostomes have been called "living fossils" in the sense that they retain many primitive features that were present in more ancient jawless fishes (e.g., ostracoderms). A recent publication, The origin and early phylogenetic history of jawed vertebrates (Brazeau, 2017) explains how an early split occurred (490 million years ago) between a group ancestor to the actual jawless fishes and a group ancestor to the jawed fishes. Soon after that (455 mya), bony and cartilaginous fish diverged. Therefore, the three groups developed parallelly and are not ancestors.



Hagfish have a skull but no vertebrae. Rudimentary vagal pathways are present in both hagfishes and lampreys. However, the vagus nerve doesn't innervate the hagfish heart and activates the lamprey heart instead of slowing it down (Taylor, 2014). Although both lack the sympathetic chains found in jawed vertebrates (gnathostomes), they have their own way of activating. The hagfish heart stores large amounts of catecholamines (Nilsson, 1998), which produces an adrenergic cardiac tone. Administration of an adrenaline antagonist causes severe bradycardia.
Gnathostomes (fishes with jaws) appeared about 450 mya as cartilaginous and bony fishes.
Cartilaginous fishes (450 mya), such as sharks, have partial vagal innervation of the heart (not the ventricles) and a robust adrenergic hormonal system. Catecholamines modulate vagal control. Sharks are fast swimmers and powerful fighters. When caught, they struggle in nets for long periods and produce large amounts of catecholamines in their blood. Acetylcholine has an excitatory effect on the ventricular muscle of the heart (Thompson, 1997).
Bony fishes are the first vertebrates to show dual heart innervation. However, we shouldn't view cyclostomes, cartilaginous, and bony fishes in a hierarchical construct. Most of them appeared in the same time window, so current living forms can't be considered the ancestors of the others.
A hagfish, a great white shark, and a bony fish. These three species do not illustrate an ascending order of dominance. Lampreys often attack other fish and even humans. In the oceans, many cartilaginous fishes can compete with aquatic mammals.
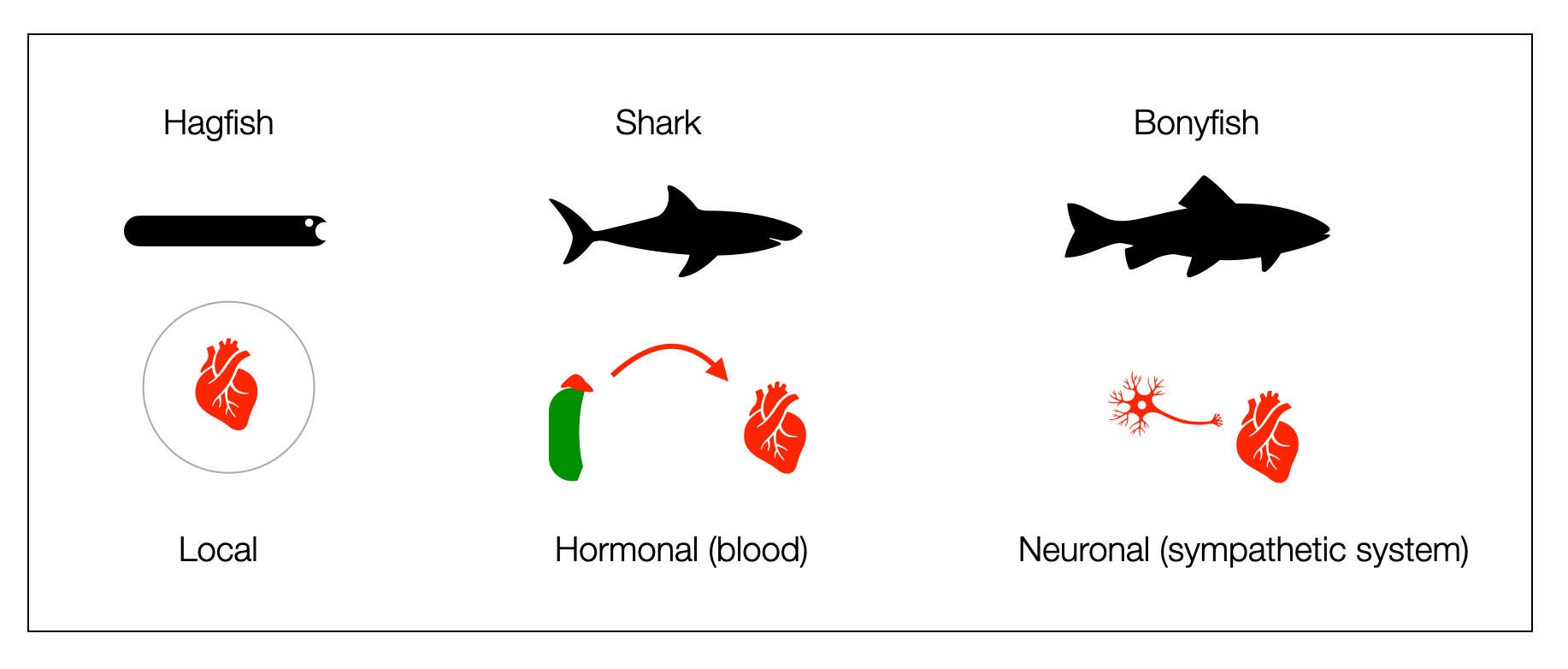
This diagram compares the adrenergic system (heart stimulation) of hagfish, shark, and bony fish. We can’t place the three groups in a hierarchical construct like a polyvagal ladder. The divergence cyclostomata (jawless) is around 490 mya, and the divergence cartilaginous and bony fish around 455 mya.
2.2.5
Evolution
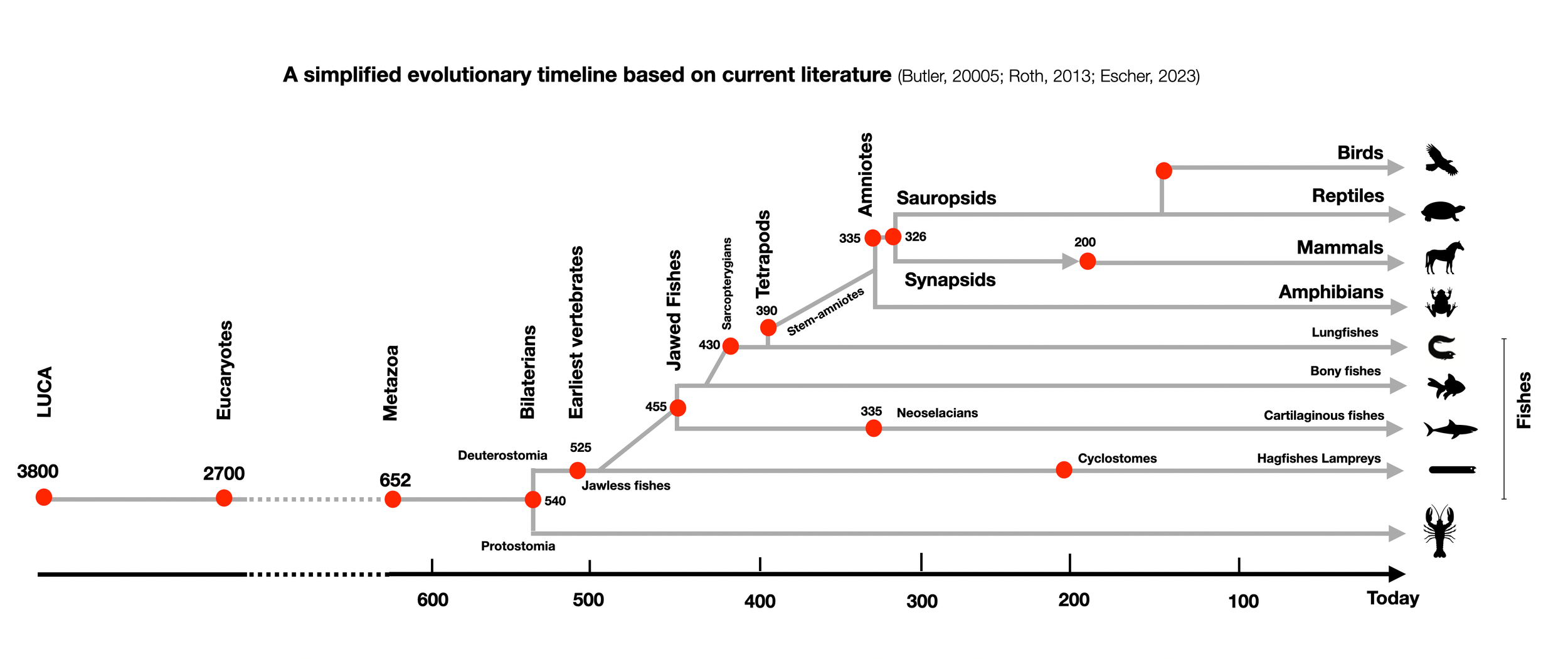
This diagram is one of the most important illustrations on this website because it directly challenges the "ladder" perspective. It shows that cartilaginous and bony fishes didn't evolve successively (ladder perspective), but in parallel (cladistic perspective). Both groups have their special cardiovascular regulation, which was successful because both survived. True cyclostomes (hagfish and lamprey) appeared relatively late-shortly before mammals. (©absb, 2024).
So-called “reptiles” are not our ancestors. We both descend from a common ancestor who lived in the short period between the appearance of amniotes and the great split between sauropsids (ancestors of “reptiles,” dinosaurs, and birds) and synapsids (ancestors of mammals).
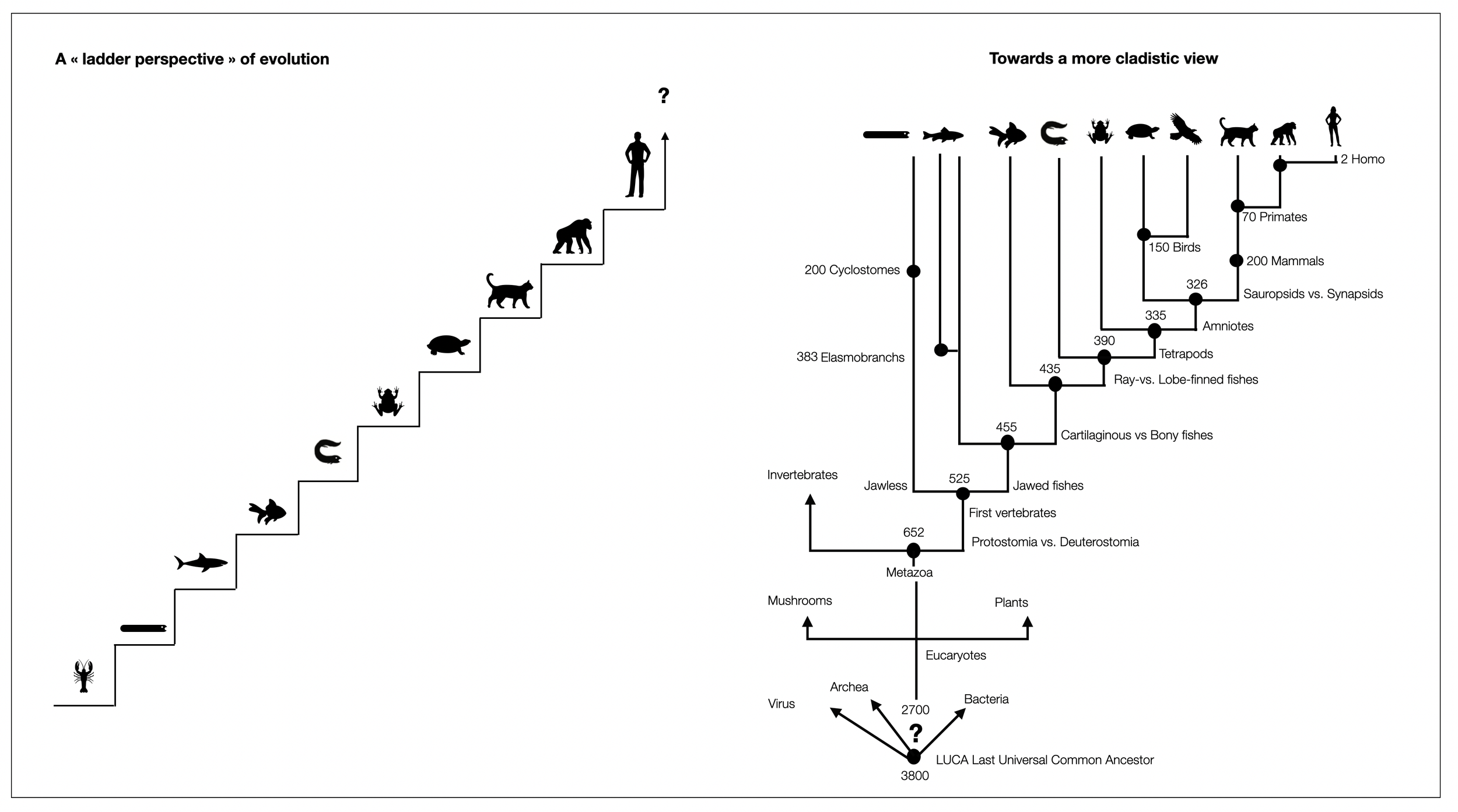
Two opposing views (diagram ©absb, 2024).
Evolution is a staircase to heaven, with each group being the ancestor of the next.
Evolution as a comb or a simplified cladogram: unlike the Olympic Games, every competitor who has survived today has an equal place at the finish line.
However, this cladogram is skewed in favor of Homo sapiens, who is on the far right.
Anthropocentrism in cladograms (Sandvik, 2009) describes this tendency in many past and recent scientific works, even textbooks that have explicitly adopted a cladistic perspective. Sandvik goes on: “One might have thought that with the acceptance of Darwinian evolution and cladistic methodology, anthropocentrism was finally banned from evolutionary narratives, at least in scientific publications (…). However, as my results show, this is merely wishful thinking. Two distorting devices can still occur even in state-of-the-art phylogenetic cladograms, and at least one of them is prevalent in textbooks: the taxon containing Homo sapiens is quite consistently placed at the rightmost position of each cladogram.”
2.2.6
Cardiorespiratory Interactions
Reading Porges gives the impression that cardiorespiratory interactions began with mammals. He prefers to speak of RSA (respiratory sinus arrhythmia) instead of HRV. RSA emphasizes the role of the ventral parasympathetic system, while HRV includes other parameters, such as the sympathetic system.
In The Phylogeny and Ontogeny of Autonomic Control of the Heart and Cardiorespiratory Interactions in Vertebrates (Taylor, 2014), the authors provide an extensive review of the development of the autonomic nervous system and, more specifically, the interactions between heartbeat and respiration. Cardio-respiratory synchrony (CRS) was found in dogfish, which was abolished by vagotomy or atropine. In addition, 45% of the CVPNs (cardiovagal neurons) are in the ventral position, while the rest are in the DVMN (dorsal). A similar organization exists in the rattlesnake (Filogonio, 2019).
The following diagram illustrates how vertebrates universally display cardio-respiratory interactions at all levels. Contrary to PVT claims, these interactions are not limited to mammals but are present in most vertebrates, perhaps for energetic reasons. In cartilaginous and bony fishes, respiratory inputs trigger the acceleration of the heartbeat, sparking further curiosity about the underlying mechanisms.
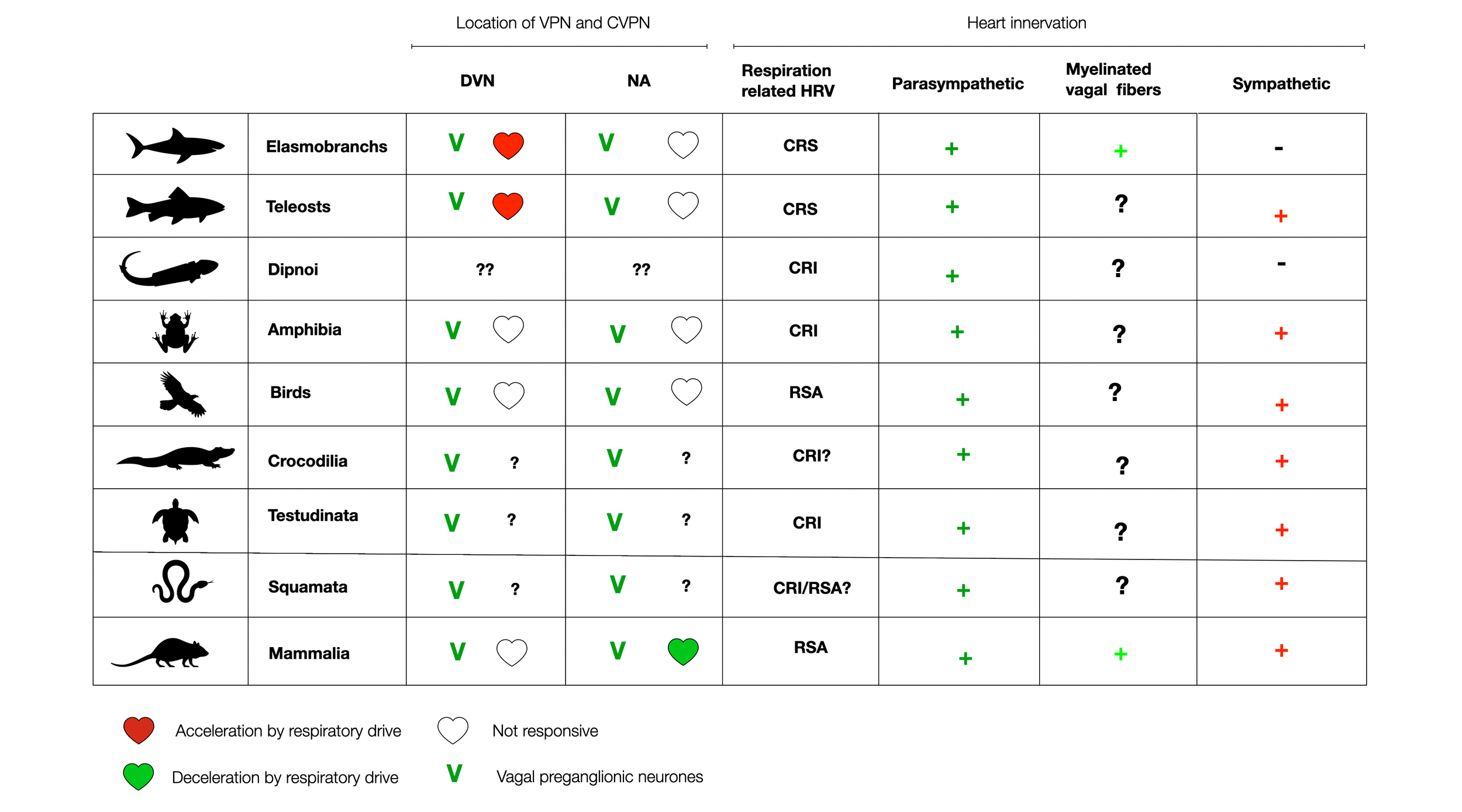
Adapted from (Taylor, 2014),(©absb, 2024). Please use the link to get access to the original article!
2.2.7
Challenging the polyvagal perspective
The current literature challenges the simplistic polyvagal phylogeny:
Ancestral jawless fish have not left enough evidence to provide sufficient information about their autonomous nervous systems. Since hagfish and lampreys (cyclostomes) appeared only 200 million years ago (mya), they cannot represent a “first” step in Dana's “evolutionary timeline” (see above).
The appearance of cartilaginous and bony fishes in the same time window (Devonian) from an ancestral jawed fish adds to the confusion. How can we find clear steps in this evolutionary process?
The question of which animal is the ancestor is complex. All three groups - cyclostomes, cartilaginous fish, and bony fish - have found a way to activate their adrenergic systems and speed up their hearts in situations of stress and challenge. Their survival to this day underscores their adaptability and resilience.
The transition from intrinsic to humoral and neural activation mechanisms is undeniably a step forward. However, it's important to note that these transitions did not occur in a precise temporal sequence, challenging the traditional view of evolution. Physically, all three mechanisms allow for similarly adrenergic solid activation.
The numbers presented by Dana (2018) (500, 400, and 200) are not based on concrete evidence, but rather seem to be arbitrary. This challenges the current understanding of the polyvagal phylogeny and encourages critical thinking about the validity of these numbers.
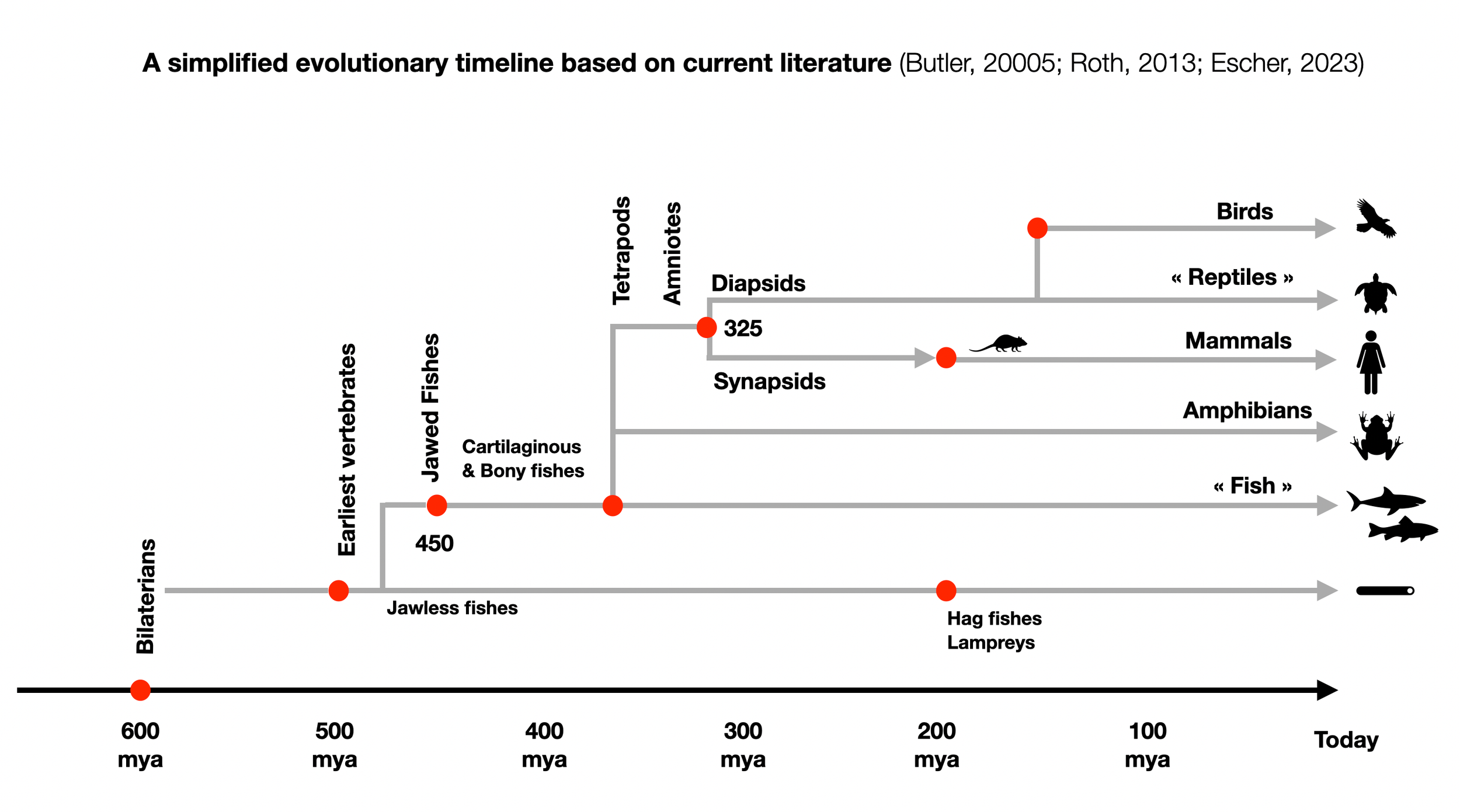
©absb, 2024
2.2.8
Cladistic vs. Polyvagal Perspective
Does the cladistic view of evolution change how we see the organization of the brainstem? Yes, it does.
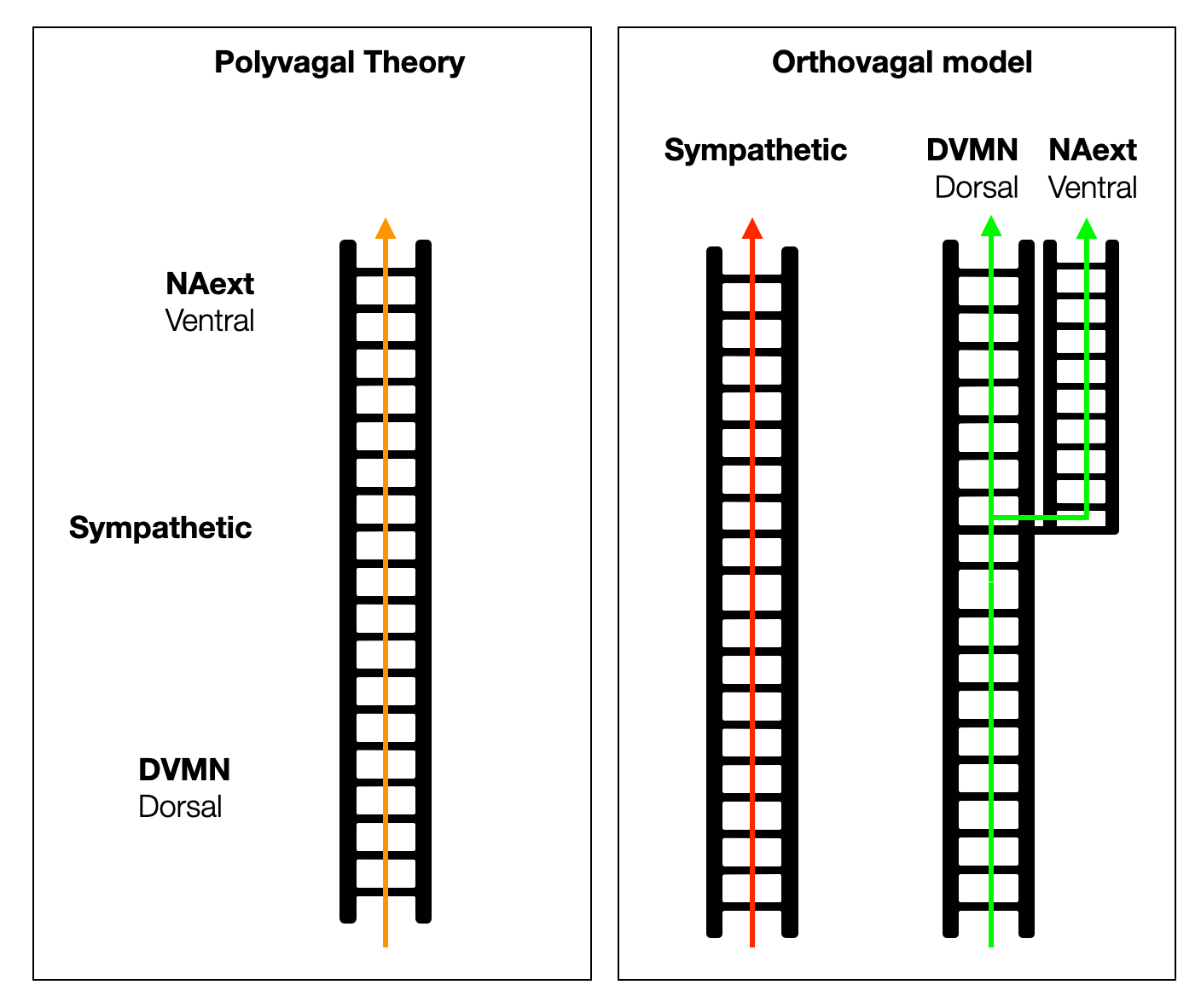
The PVT aligns the dorsal, sympathetic, and ventral systems on a unique vertical ladder as if they evolved in a straight line, one on top of the other. In reality, the sympathetic and parasympathetic systems developed in parallel from an ancient organization to a high level of complexity. The adrenergic system first relied on intrinsic cardiac catecholamines, then on circulating (hormonal) catecholamines, and finally took physical form as an anatomical system.
From hagfish to birds, the parasympathetic system controls all parasympathetic functions with one dorsal branch. In mammals, the slowing of the heartbeat was moved to a ventral position in the NAext, creating a new branch. However, a small proportion of cardiac vagal neurons (CVPNs) remain in the DVMN to protect and support the left ventricle. The classification of the DVC and the VVC (an artificial construct) as “old” and “new” is not accurate. The DVC is the central manager of visceral life — what should be “old” there?
Therefore, we can draw a simplified model with two or even three ladders instead of a single one, knowing that a ladder is always a dangerous metaphor.
2.2.9
Embryology
During the first few weeks of life, the embryo develops into a crescent-shaped structure with folds at the front: the branchial or pharyngeal arches, the remains of the gills in fish.
Looking at the upper part of the embryo, we see an invagination of the ectoderm, giving birth to an elongated structure called the neural tube, which becomes the central nervous system (brain and spine).
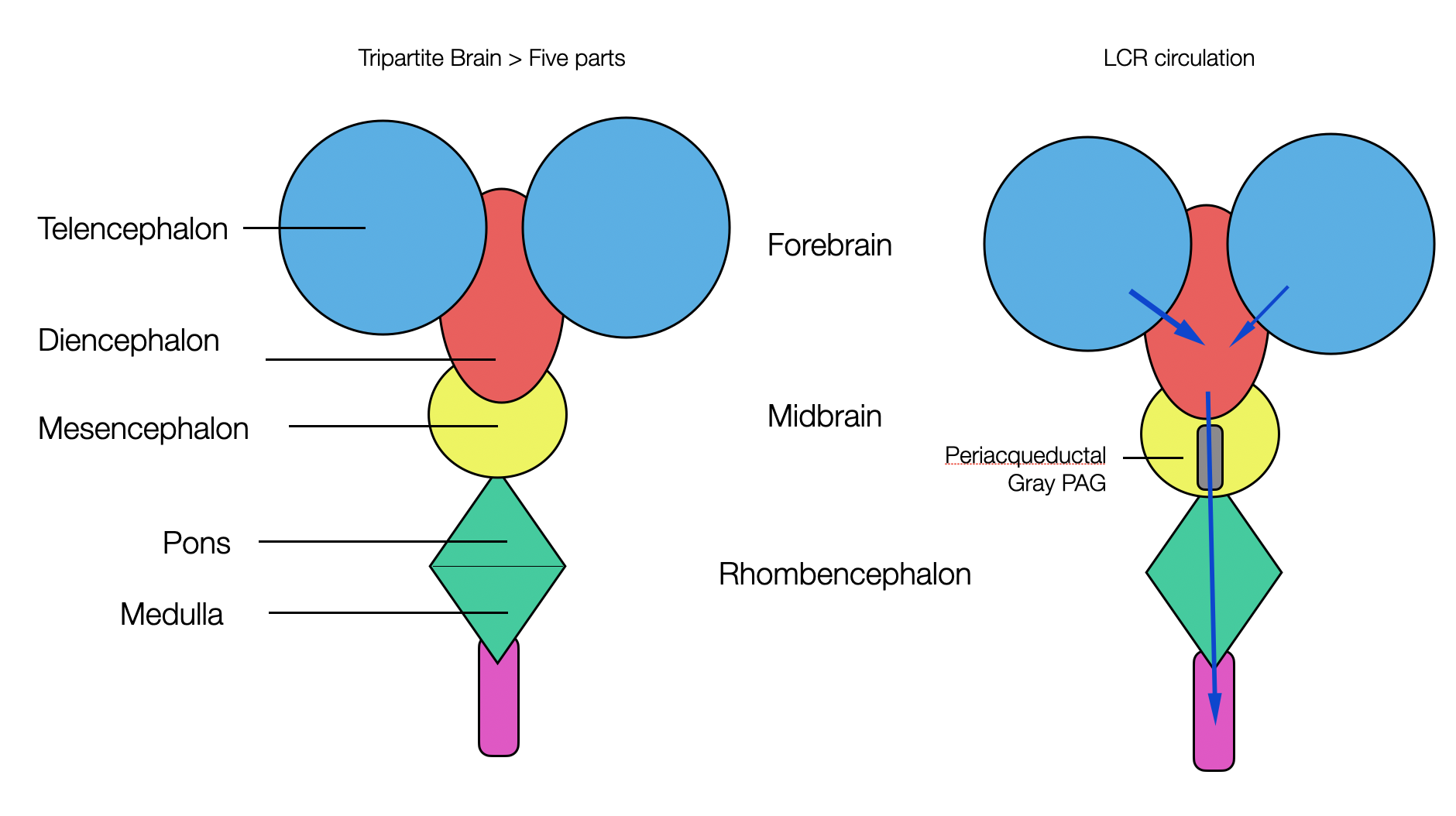
All vertebrates have a brain consisting of three parts:
The forebrain (prosencephalon), which later divides into the telencephalon and diencephalon.
Middle brain (mesencephalon)
The hindbrain (rhombencephalon) later divides into the metencephalon (or pons) and the myelencephalon (or medulla).
©absb, 2024
The chambers develop into the first, second, third, and fourth ventricles.
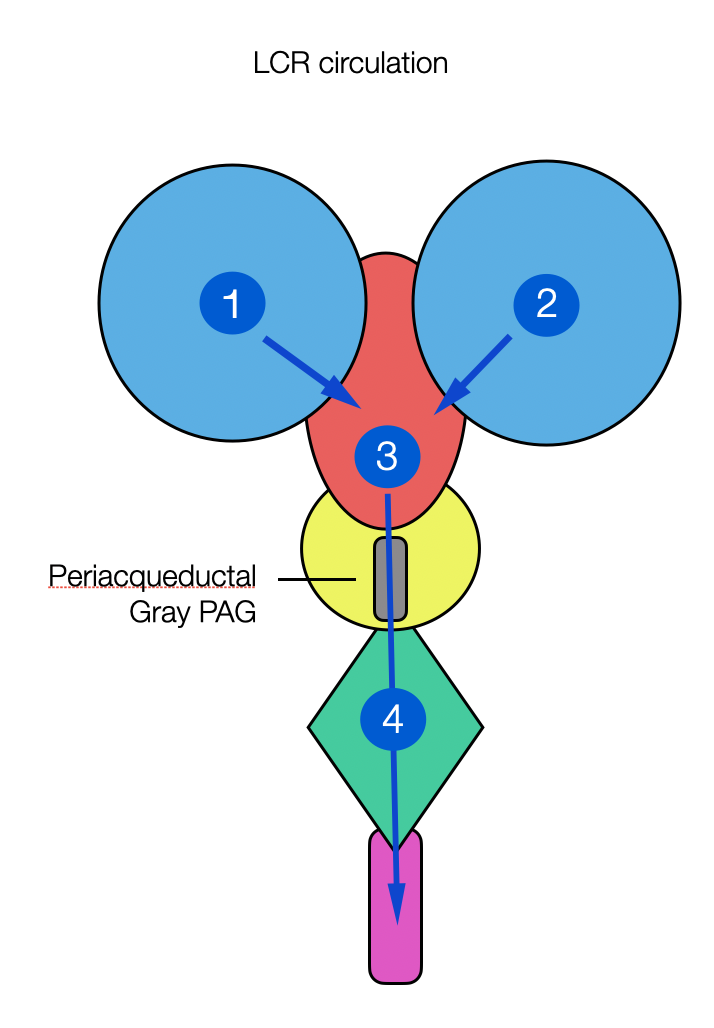
A narrow channel called the aqueduct connects the third and fourth ventricles. Around it will later appear a shaft of gray matter called the periaqueductal gray (or PAG). This is where the fight-flight-freeze responses originate.
2.2.10
Branchial - Pharyngeal Apparatus
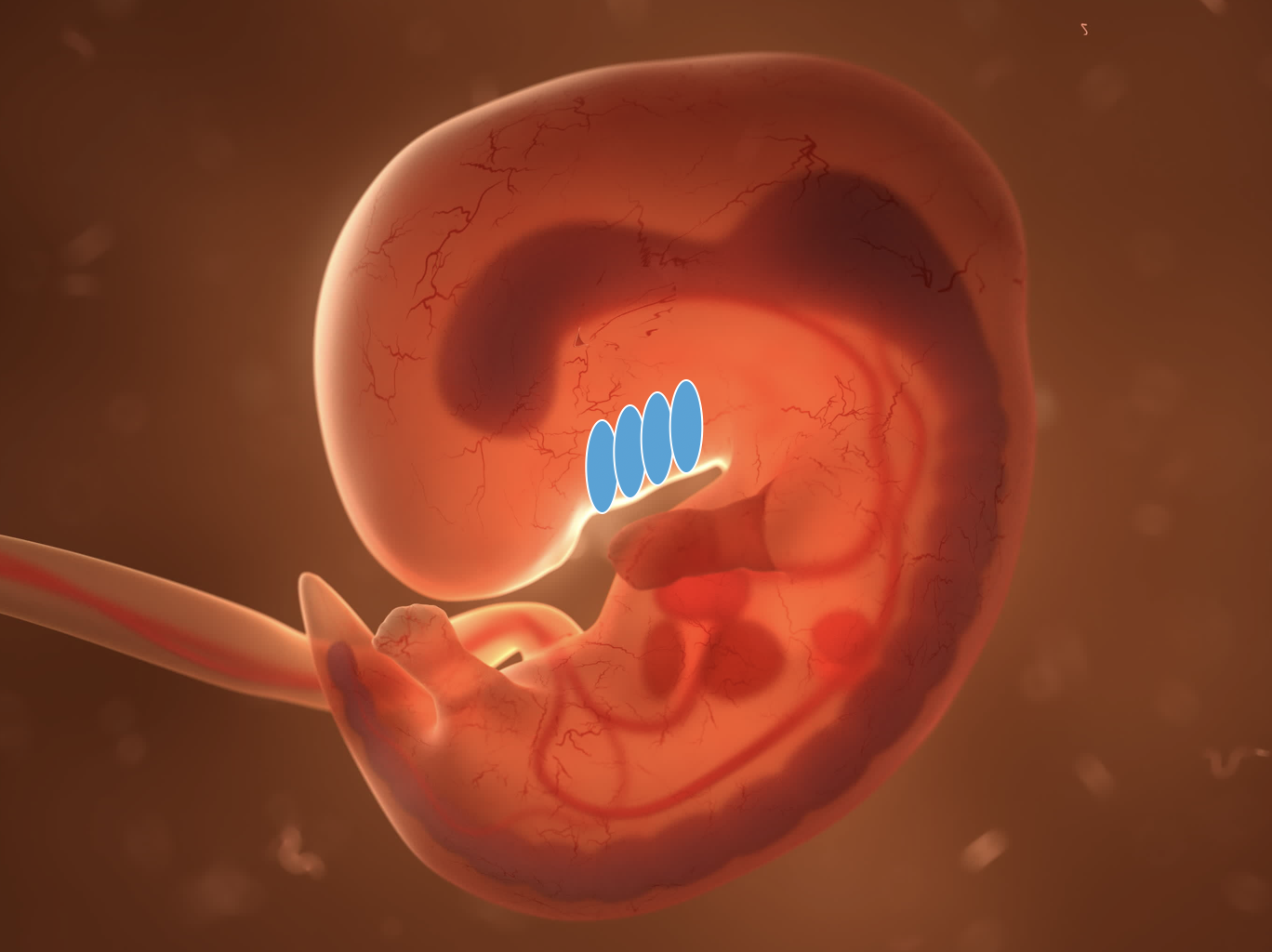
In the first weeks of embryonic development, folds appear in the neck. Similar pouches appear in the pharynx. Together, they form arches that are innervated by the branchiomotor (or pharyngeomotor) cranial nerves V, VII, IX, and X. However, they don't develop into gills as in fish — that would be recapitulation.
Vertebrate embryo. The blue ovals indicate where the branchial arches appear.

Branchial arches: graphic by Wikipedia.

Grey nurse shark showing branchial arches (gills)—source Wikimedia.
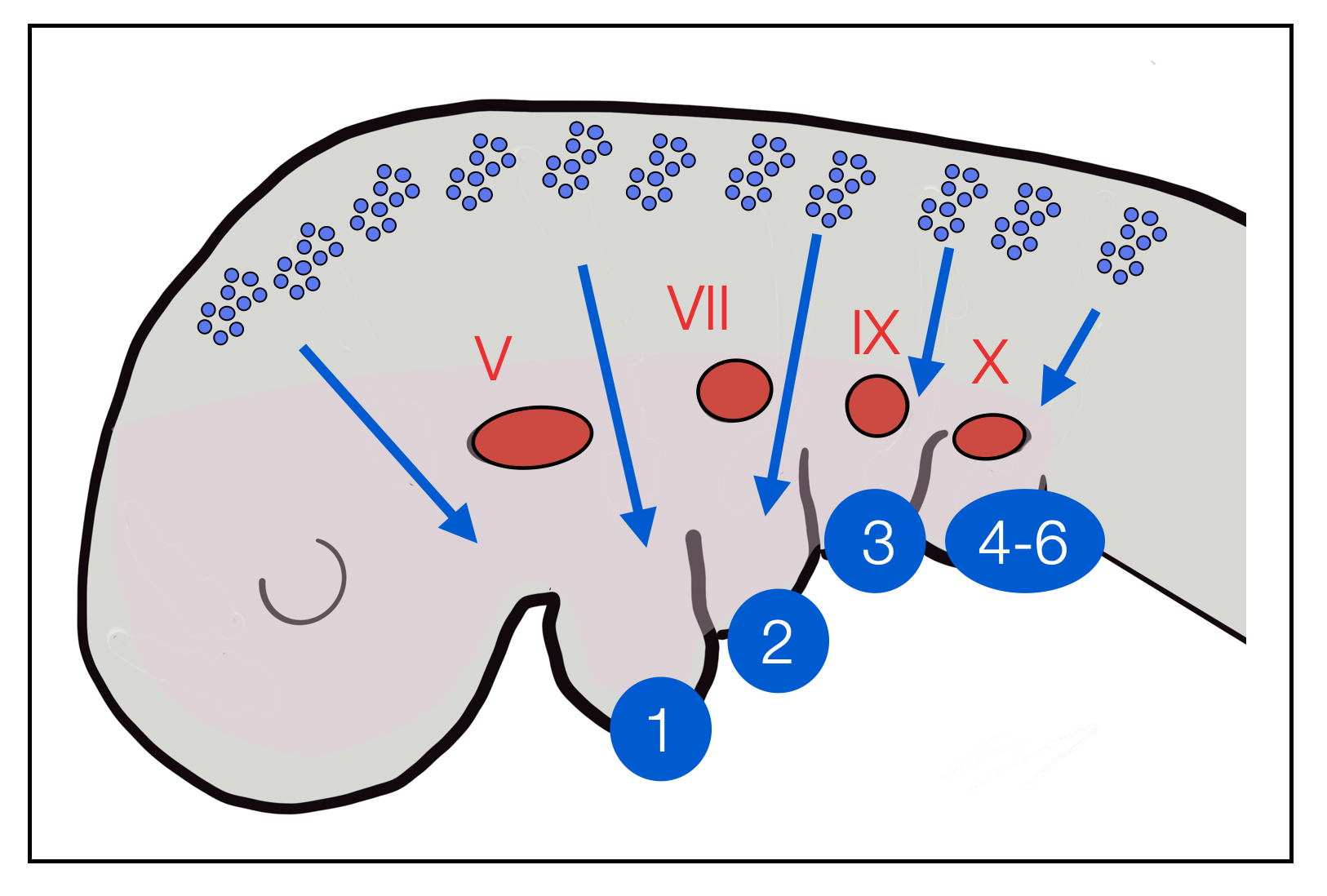
Vertebrate embryo. From the neural crest in the head region, cells migrate ventrally (pharyngeal arches) to form structures of the face and neck. Blue numbers: pharyngeal (branchial) arches. In red: nuclei of motor cranial nerves V, VII, IX and X + epipharyngeal placodes. (see Neurogenic placodes on Wikipedia).
2.2.11
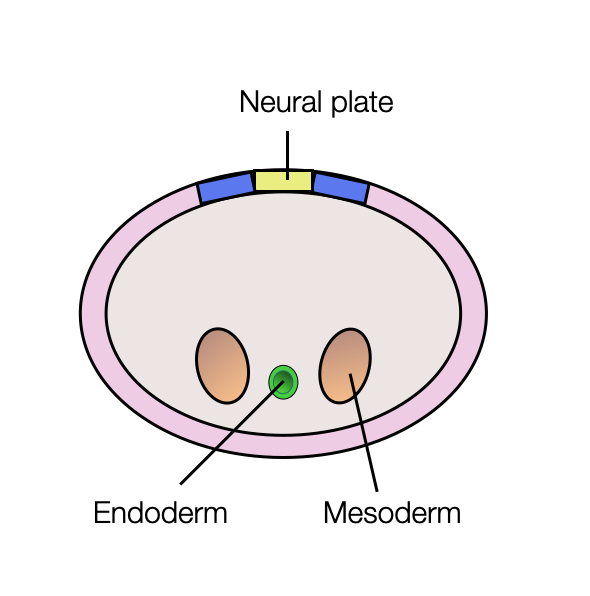
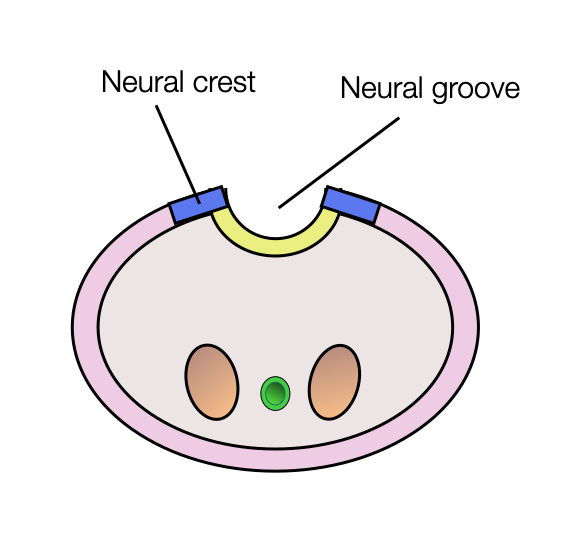
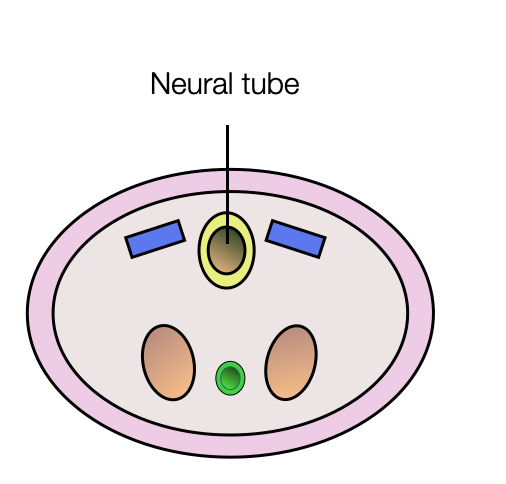
Neural Tube and Crests
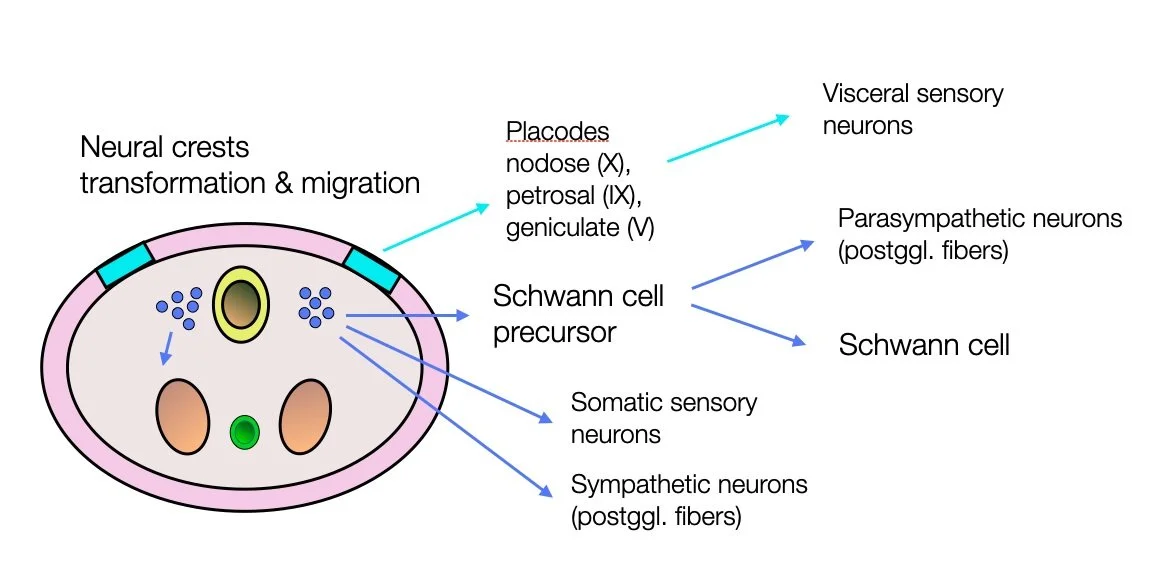
The neural tube is formed by invagination of the ectoderm (neural plate). In parallel, two formations appear on each side of the tube — the neural crests. They transform into multipotent cells that migrate to different regions of the embryo. They differentiate into a wide range of cell types, including peripheral and enteric neurons, glia, melanocytes, and smooth muscle. In the cranial region, these cells contribute to most of the cartilage and bone of the skull and the pharyngeal skeleton.
During embryonic development, several migrations take place. After the neural crests have switched from an epithelial to a mesenchymal organization, they become stem cells and give rise to various cell types: melanocytes, craniofacial cartilage and bone, smooth muscle, peripheral and enteric neurons, and ganglia. These new cells migrate in different directions. At the head and neck level, they contribute to forming the branchial arches — Source: Establishing neural crest identity: a gene regulatory recipe (Simões-Costa, 2015).

2.2.12
Origins of the Central & Peripheral Nervous System
The central nervous system (CNS) arises from the neural tube (shown in yellow).
The peripheral nervous system (PNS) arises from the neural crests and placodes.
Sympathetic and parasympathetic postganglionic fibers have the same origin (neural crests).
2.2.13
Distribution of Nuclei in the Brainstem
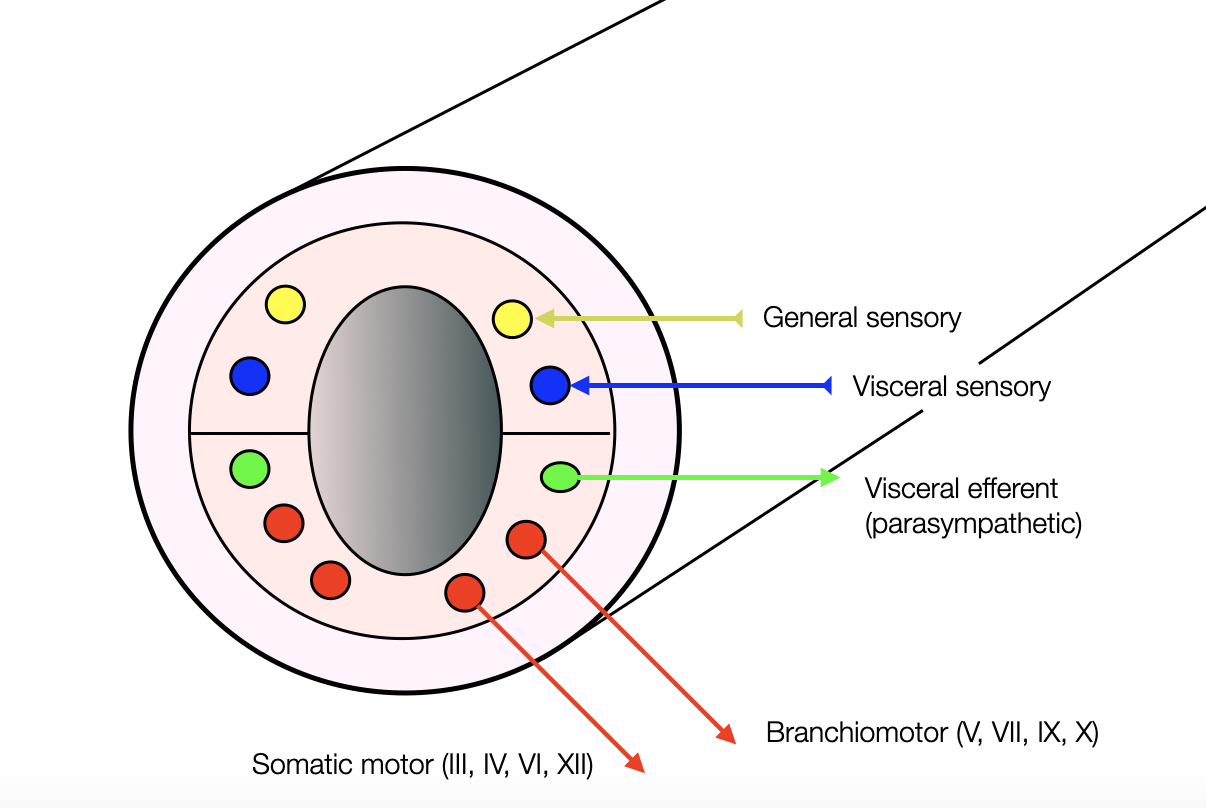
Distribution of the nuclei in the neural tube (first weeks of development): The sensory nuclei are in the upper part, and the motor nuclei are in the lower part.
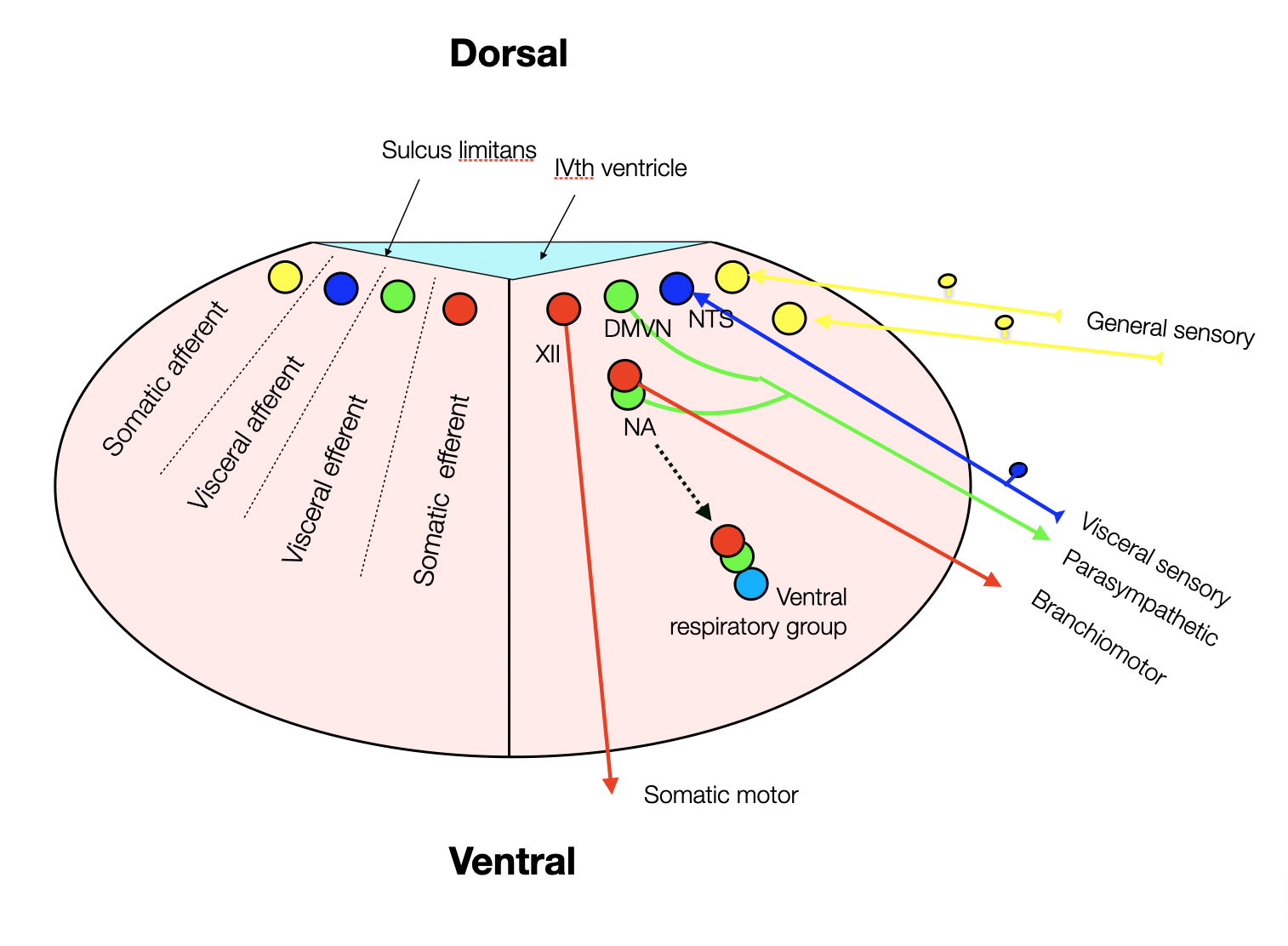
During embryonic development, the nuclei evolve into a more horizontal arrangement as the fourth ventricle becomes flatter. The NA (nucleus ambiguus), which has motor and visceral parts, migrates to a ventral lateral position just above the ventral respiratory group.
2.2.14
Ontogeny and PVT
Embryonic & fetal development
The PVT proposes embryonic and fetal development in three stages, similar to the three-stage development from DVC to sympathetic and VVC. The problem is that the VVC amalgam has too many heteroclite elements, making the discussion difficult:
Branch motor nerves (V, VII, IX, X, and XI) control striated muscles like any other somatic motor nerve. We can use them at will, even if we don't understand the complexity of their coordination. They don't have to be fully myelinated to be functional. However, the full maturation of the nervous system continues after birth.
The cardiac vagal neurons (CPVNs), which form the ventral vagal (parasympathetic) branch, migrate with the vagal neurons from a dorsal to a ventral position. Like the branchiomotor nerves, the ventral vagal branch is functional before migration and complete myelination, which continues after birth.
Myelinization accelerates neuronal speed. Nerves that rely on this improvement are more efficient, but also more vulnerable. Any delay in myelination can cause serious problems (e.g., SID). Non-myelinated pathways are slower but more stable. So, let's not compare apples and oranges.
The polyvagal three-step description of ANS development is not convincing.
In the chapter about Heart Rate Variability (HRV), we dedicated a section to the Fetal HRV (fHRV), which doesn’t support the idea of hierarchy of the fHRV.
Further Reading
Central vagal sensory and motor connections: human embryonic and fetal development (Cheng, 2004).
Development of the human dorsal nucleus of the vagus (Cheng, 2008).
The Initial Appearance of the Cranial Nerves and Related Neuronal Migration in Staged Human Embryos (Müller, 2011). By stage 16, all proper cranial nerves are recognizable. In stages 13-18, neuronal migration is a characteristic feature in the development of all cranial nerves except the somatic efferent nerves (III, IV, VI, XII). The authors don't mention any difference in development at any stage.
Fetal Development of the Central Nervous System (Ashwell, 2012) He gives a detailed report on the fetal development of the cranial nerve nuclei. Despite repeated readings, we couldn't find valuable arguments for a three-stage development in the sense of the PVT. Most major motor nuclei (V, VI, X, and XI) reach their final positions at 8 w.g. (= week of gestation). The rest need another two weeks. At 9 weeks' gestation, the DVMN has not yet reached its final position. At 13wg, a distinct structure appears, but maturation continues until birth. All adult subnuclei of the DVMN appear by 21 w.g. and are essentially adult by 25 w.g. The connection of the NTS to the DVMN is not well-developed until 24 w.g. The structural development of the DVMN appears to occur in parallel with the functional maturation of cardiovascular and gastric movements (Cheng, 2008). Insufficient maturation of respiratory centers in the human fetal brainstem may cause Sudden Infant Death (SID). Since most involved structures (NTS, respiratory centers) mature in the third trimester (Lavezzi, 2004), premature infants are particularly vulnerable.
>> to the next chapter Hierarchy